 |
|
МЕНЮ
|
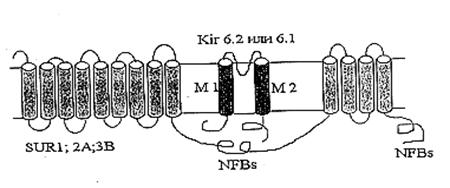
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию1.2 АТФ-зависимый калиевый канал цитоплазматической мембраны К настоящему времени КАТФ каналах плазматической мембраны хорошо изучены. Изначально их классификация велась по величине проводимости одиночных каналов [Ashcroft and Ashcroft, 1990], а позднее, с развитием молекулярно-биологических методик, по аминокислотному составу белковой молекулы канала [Yokoshiki et al., 1998]. 1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала Молекулярно-биологические и электрофизиологические исследования последних лет позволили определить структуру цитоКАТФ канала. Исследуемый канал состоит из двух белков: KIR – inward rectifying K+ channels, формирующий пору канала, и SUR – sulphonyl urea receptor, регуляторная субъединица, придающая каналу чувствительность к модуляторам. SUR-субъединица содержит нуклеотидсвязывающие участки, локализующиеся на цитоплазматической стороне, а также участки связывающие активаторы калиевых каналов (potassium channel activators, PCOs) и MgАДФ. Каналы подсемейства KIR6.0 сами обладают чувствительностью к АТФ. Роль SUR заключается, вероятно, в облегчении доступа АТФ к АТФ-связывающему участку KIR [Tucker et al., 1997; Yokoshiki et al., 1998]. В настоящее время механизмы молекулярного взаимодействия KIR и SUR окончательно не выяснены. Результаты исследования электрофизиологических свойств нативных КАТФ каналов позволяют предположить, что они принадлежат к классу калиевых каналов внутреннего выпрямления (KIR). В 1993 году на многие KIR каналы (KIR1.1 [Ho et al., 1993], KIR2.1 [Kubo et al., 1993], KIR3.1 [Dascal et al.,1993]) были получены клоны. Их выделили в отдельное подсемейство KIR-каналов (KIR1.0-6.0), основанное на унифицированной номенклатуре [Chandy and Gutman, 1993]. Гидрофобные участки клонированных KIR каналов содержат два мембранопересекающих сегмента М1 и М2, образующих пору. Подобно потенциалзависимым К+ каналам, KIR каналы, вероятно, формируют тетрамер [Doupnik et al., 1995; Salkoff and Jegla, 1995; Nichols et al., 1996; Nichols and Lopatin, 1997]. Также был клоирован регуляторный белок, содержащий участок, связывающий сульфонилочевину (SU-связывающий сайт) [Aguillar-Bryan et al., 1995; Thomas et al., 1995; Inagaki et al., 1996; Isomoto et al., 1996]. Клонированные регуляторные белки получили название SUR. SUR содержит два нуклеотидсвязывающих участка (NFBs) с цитоплазматической стороны. N-конец находится с внешней стороны мембраны и содержит два домена, предположительно из 9 и 4 пронизывающих мембрану спиралей (Рис.2) [Yokoshiki et al., 1998].
Рис. 2. Схематическое изображение цитоплазматического КАТФ канала как комплекса KIR и SUR [Yokoshiki H. et al., 1998] Считается, что нативный КАТФ канал представляет собой комплекс из четырех KIR6.x и четырех SUR субъединиц [Clement et al., 1997]. Различные типы регуляторных субъединиц, SUR1, SUR2A и SUR2B, обнаружены, соответственно, в поджелудочной железе, сердце и гладких мышцах [Ammala et al., 1996; Inagaki et al., 1996; Yamada et al., 1997]. 1.3 АТФ-чувствительный калиевый канал внутренней мембраны МХ В 1981 г. из внутренней мембраны МХ был выделен К+-селективный канал (митоКАТФ) [Миронова и др., 1981], который ингибируется физиологическими концентрациями АТФ [Paucek et al., 1992; Миронова и др., 1996 (I)]. В 1991 г. этот канал был впервые обнаружен во внутренней мембране МХ методом пэтч-кламп [Inoue et al., 1991], позднее появились аналогичные работы других исследователей [Nakae et al., 2003; Fikret et al., 2004; Dahlem et al., 2004; Bednarczyk et al., 2005]. В настоящее время канал активно изучается, поскольку обнаружено, что он участвует в адаптации организма к гипоксии [Garlid et al., 1997]. В настоящее время основными методами выделения АТФ-ингибируемого МХ калиевого канала (митоКАТФ) является метод водно-этанольной экстракции [Миронова и др., 1981; Миронова и др., 1996 (I)] и солюбилизация белка-канала из мембраны Тритоном Х-100 с последующей очисткой ионообменной хроматографией. При этом митоКАТФ канал выделяют в виде белка с молекулярной массой 55 кДа [Mironova et al., 2004]. 1.3.1 Структурная организация митоКАТФ канала Так как митоКАТФ имеет ряд общих свойств с цитоКАТФ и регулируется общими регуляторами, было предположено, что по структуре он близок к цитоплазматическому каналу [Mironova et al., 1999] и состоит из канальной субъединицы [Mironova et al., 2004] и регуляторной [Bajgar et al., 2001]. Белок с м. м. 55 кДа, выделенный из внутренней мембраны МХ печени крысы, является, по-видимому, канальной субъединицей митоКАТФ. При встраивании в БЛМ этот белок формирует АТФ-зависимые каналы (митоKIR). Активность этих каналов не блокируется глибенкламидом и специфическим ингибитором митоКАТФ 5-ГД, и не активируется кромакалимом и ддиазоксидом. Однако эти препараты влияют на АТФ-зависимый калиевый транспорт в изолированных МХ, где есть обе субъединицы канала [Mironova et al., 2004]. МитоКАТФ, также как и клеточный, является, по всей вероятности, гетеромультимером, состоящим из калиевого канала, белка с молекулярной масой 55 кДа, который имеет выпрямляющие свойства и который, по аналогии с цитоплазматическим каналом, был назван митоKIR (от англ. Inward rectifying) [Mironova et al., 2004] и рецептора, чувствительного к сульфонилмочевинам и поэтому названного митоSUR (от англ. sulphonylurea) (Рис. 3) [Mironova et al., 1999; Bajgar et al., 2001]. Кi для АТФ на реконструированном в БЛМ митоKIR выше, чем в интактных МХ и Mg2+ для этого ингибирования не требуется. Тогда как для АТФ-ингибирования канала, локализованного в интактных МХ, присутствие магния было необходимо [Mironova et al., 2004]. Те же различия были обнаружены при сравнительном изучении влияния АТФ и сульфонилмочевин на цитоKIR и целый цитоКАТФ [Tucker et al., 1997]. Известно, что основной участок связывания АТФ в цитоКАТФ локализован на канальной субъединице. Регуляторная субъединица повышает сродство канальной субъединицы к АТФ и обеспечивает чувствительность целого канала к активаторам и ингибиторам [Tucker et al., 1997].
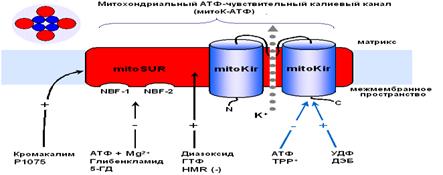
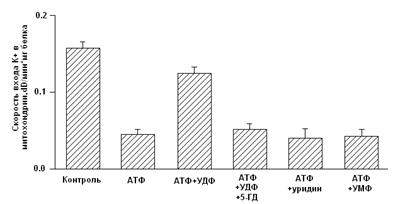
Рис.3. Рабочая модель структуры МХ АТФ-зависимого К+ канала [Mironova et al., 2004] Приведенные выше результаты исследований митоКАТФ подтверждают предположение о том, что белок с м.м. 55 кДа является канальной субъединицей целого митоКАТФ. Функцию регуляторной субъединицы выполняет, вероятно, белок с м.м. 63 кДа, связывающийся с меченым глибенкламидом [Bajgar et al., 2001]. В то же время, результаты исследований, проведенных на МХ сердца, с использованием антител на Kir6.1 и Kir6.2, а также на SUR1 и SUR2, показали, что в МХ кардиомиоцитов присутствуют Kir6.1 и Kir6.2, но нет ни SUR1, ни SUR2. Однако в них обнаруживается SUR2-подобный белок, который, возможно, является сплайсированным вариантом SUR2 [Lacza et al., 2003 (I)]. При исследовании митоКАТФ мозга, близкого по структуре к цитоКАТФ [Lacza et al., 2003 (II)], с использованием АТ к KIR6.1 и KIR6.2, а также SUR1 и SUR2 субъединицам цитоКАТФ было показано, что очищенные МХ мозга содержат преимущественно KIR6.1 и, в меньшей степени, KIR6.2 и не содержат SUR1 и SUR2. Согласно выводам авторов, в формировании митоКАТФ мозга участвует укороченный вариант SUR2 или белок, подобный по структуре SUR2. Кроме того, показано, что МХ мозга содержат в 8 раз больше KIR6.1 и SUR2-подобного белка, по сравнению с общим белком ткани мозга, что подтверждает локализацию этих белков в МХ [Lacza et al., 2003 (II)]. Тот факт, что м.м. митоKIR мозга несколько ниже (50 кДа), чем у митоKIR печени [Mironova et al., 1999; 2004], предполагает наличие тканевой специфичности в структуре каналов. Было показано, что при встраивании в липосомы или липидные бислойные мембраны комплекса митохондриальных белков, состоящего из АТФ связывающего кассетного белка-1 (mАВС1 от англ. Mitochondrial binding casset), АТФ-азы, сукцинатдегидрогеназы, аденилаттранслоказы и фосфатного переносчика, в мембране образуются чувствительные к АТФ и глибенкламиду каналы [Ardehali et al., 2004]. Это послужило основой для другого варианта модели субъединичной организации митоКАТФ [Ardehali and O'Rourke, 2005]. Однако нельзя исключить наличие в этом белковом комплексе других митохондриальных белков, включая митоKIR, тем более что белок с м.м. 55 кДа в нём присутствует. Поcкольку этот мультикомплекс в липосомах индуцировал открытие неспецифической Са2+-зависимой поры в присутствие ионов Са2+ [Ardehali et al., 2004], это говорит о возможном загрязнении исследуемого комплекса порином и другими белками. Было показано, что один из белков этого комплекса, а именно mАВС1, играет важную роль в защите клеток миокарда от окислительного стресса [Ardehali et al., 2005]. Однако кардиопротекторное действие этого белка не снималось ингибиторами митоКАТФ - глибенкламидом и 5-НD. Поэтому, по мнению авторов, пока не получено достаточно доказательств для утверждения того, что mАВС1, имеющий, также как и митоКIR, м.м. 55 кДа, является частью митоКАТФ. Таким образом, вопрос структуры митоКАТФ по-прежнему остается открытым. В связи с чем, в представленной работе была предпринята попытка определить относится ли белок с м.м. 55 кДа, выделяемый из внутренней мембраны МХ методом водно-этанольной экстракции, к системе АТФ-зависимого транспорта К+. Также, был проведен анализ гомологии структуры исследуемого белка последовательностям известных белков. Глава 2. Модуляторы 2.1 Модуляторы митоКАТФ канала 2.1.1 Метаболические модуляторы митоКАТФ канала В связи с выраженным кардиопротекторным действием активаторов митоКАТФ в настоящее время во многих лабораториях ведется поиск новых синтетических активаторов канала, которые могут быть потенциальными кардиопротекторами [Ockaili et al., 2002; Kopustinskene et al., 2001]. В нашей лаборатории помимо синтетического донора электронов ДЕБ, который селективно активировал митоКАТФ [Mironova et al., 1999; Mironova et al., 2004], был обнаружен ряд природных, то есть метаболических активаторов этого канала. Дифосфонуклеотиды Показано, что дифосфонуклеотиды (АДФ и ГДФ) активируют реконструированную в БЛМ канальную субъединицу (Рис. 2) [Grigoriev et al., 1999; Mironova et al., 1999]. Данные по активирующему действию ГДФ в дальнейшем были подтверждены и другими исследователями [Bednarczyk et al., 2005]. Недавно в нашей лаборатории было показано, что в ряду дифосфонуклеотидов наиболее эффективным является уридиндифосфат (УДФ) [Mironova et al., 2004]. В МХ УДФ (~20 мкМ) активирует митоКАТФ, и эта активация снимается глибенкламидом и 5-ГД (Рис. 4) [Mironova et al., 2004]. Концентрации УДФ, неоходимые для активации реконструированного в БЛМ митоKIR и целого канала в нативных МХ, одинаковы. Следовательно, участок связывания УДФ локализуется на канальной субъединице. Для активации цитоКАТФ нужны значительно большие концентрации этого препарата [Alekseev et al., 1998].
Рисунок 4. Активация УДФ целого митоКАТФ канала в нативных МХ [Mironova et al., 2004] Ранее на моделях тотальной и регионарной ишемии изолированных перфузируемых сердец крыс было показано, что уридин и УМФ проявляют антиаритмическую активность в отношении реперфузионных аритмий, предупреждают развитие миокардиального станинга при постишемической реперфузии, препятствуя депрессии сократительной функции миокарда и предотвращая развитие контрактуры [Eliseev et al., 1989]. В представленной работе была предпринята попытка определить, обладают ли уридиновые нуклеотиды кардиопротекторным действием и связано ли их влияние с активацией цито- и митоКАТФ каналов на модели искусственного острого инфаркта миокарда. 2.2 Функциональная роль митоКАТФ Являясь одним из компонентов калиевого цикла МХ, митоКАТФ участвует в поддержании объема МХ матрикса и мембранного потенциала [Garlid, 1998]. В лаборатории проф. Мироновой показано, что активация митоКАТФ играет важную роль при выходе животных из состояния зимней спячки [Fedotcheva et al., 1985; Миронова и др., 1986]. МитоКАТФ участвует также в ингибировании процесса апоптоза клетки, блокируя высвобождение цитохрома С, предотвращая снижение мембранного потенциала и чрезмерное увеличение концентрации Ca2+, сохраняя, таким образом, функции МХ, что было показано на b-клетках поджелудочной железы [Akao et al., 2001], а затем и на церебральных нейронах [Teshima et al., 2003]. Кроме того, митоКАТФ является посредником в процессе регуляции транскрипции некоторых генов. Его активация вызывает небольшое увеличение продукции активных форм кислорода (АФК) [Chandel et al., 1998]. При патологии активация митоКАТФ частично востанавливает мембранный потенциал, стимулируя дальнейший перенос протонов, ингибируя накопление внутримитохондриального кальция и предотвращая распад энергетических фосфатов путем создания оптимального для синтеза АТФ электрохимического градиента [Szewczyk, 1996; Xu et al., 2001]. Активация митоКАТФ способствует разобщению электронного транспорта, ускоряя, таким образом, дыхание, что приводит к окислению проксимальных переносчиков электронов дыхательной цепи [Liu et al., 1998]. Согласно данным Гарлида, активация канала приводит к набуханию МХ, что способствует поддержанию архитектуры внутренней мембраны и сохранению активности дыхательной цепи. Это позволяет поддерживать уровень клеточного АТФ и, таким образом, способствует восстановлению функций миокарда во время реперфузии [Garlid, 2000]. Как было показано ранее, набухание матрикса увеличивает скорость окислительного метаболизма, активирует как окисление жирных кислот, так и дыхание и синтез АТФ [Halestrap, 1989]. 2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксииВ 1983 г. был открыт феномен прерывистой гипоксии, важную роль в котором играет активация митоКАТФ [Murry et al., 1983; Liu et al., 1998]. Феномен прерывистой гипоксии был назван «preconditioning» или «ишемической толерантностью» [Murry et al., 1986; Kitigawa, 1990]. Позднее он получил название «феномен ишемической адаптации» (ФИА) [Yellon et al., 1998; Петрищев и др., 2001], и в настоящее время признан наиболее эффективным способом уменьшения размера инфаркта миокарда [Kloner et al., 1998]. Справедливости ради надо отметить, что адаптация, создаваемая кратковременной гипоксией, была обнаружена ранее [Poupa et al.,1966; Meerson et al., 1973]. Клеточные механизмы ФИА в настоящее время широко изучаются. Считается, что защитный эффект является трехступенчатым и включает в себя последовательные триггерные и конечные эффекторные механизмы, причем наиболее вероятным эффектором является митоКАТФ [Garlid et al., 1997; Yellon et al., 1998; Grover and Garlid., 2000; Петрищев и др., 2001]. ФИА развивается в виде двух волн: первая волна проявляется в течение нескольких часов, затем исчезает и снова появляется в виде второй волны [Yellon et al., 1998], которая длится в течение недели, а иногда и месяца [Carroll et al., 2000]. Установлено, что во второй стадии адаптации к гипоксии также участвует митоКАТФ [Wang et al., 2001]. В развитии толерантности к ишемии мозга [Lacza et al., 2003], и в дистанционной ишемической адаптации сердца к гипоксии при создании коротких эпизодов окклюзии почечной артерии также задействован митоКАТФ [Pell et al., 1998]. Установлено, что специфические модуляторы митоКАТФ (см. таблицу 1) влияют на ФИА. Так активатор митоКАТФ, диазоксид, и неселективный активатор КАТФ каналов, кромакалим, имитируют «preconditioning», увеличивая время сокращения миокарда и способствовуя восстановлению функций изолированных сердец крыс, подвергавшихся общей ишемии миокарда с последующей реперфузией. Кi диазоксида для митоКАТФ (0.8 мкМ в МХ сердца быка) в 1000 раз превышала Кi для цитоКАТФ [Gross et al., 1992]. Кромакалим проявлял аналогичный эффект в тех же концентрациях. Глибенкламид и 5-ГД блокировали действие диазоксида и кромакалима, следовательно, эффект активаторов на ФИА опосредован через КАТФ каналы [Auchampach, 1992; Garlid, 1997; Tsai et al., 1998; Vanden Hoek, 2000]. Активирующе действие диазоксида на митоКАТФ показано также на изолированных МХ сердца крысы, что доказывает роль МХ канала в кардиопротекции [Gross et al., 1992]. Полученные данные подтверждают также результаты исследований, по изучению флуоресценции флавопротеинов на изолированных кардиомиоцитах желудочков сердца кроликов [Liu et al., 1998; Sato et al., 1998]. Однако поскольку было обнаружено, что диазоксид снижает мембранный потенциал на МХ, независимо от активации митоКАТФ канала, его кардиопротекторный эффект вызывает сомнение. Показано, что потенциальный медиатор сигнального пути, ответственного за формирование ишемической адаптации, протеинкиназа С, [Sato et al., 1998; Liu et al., 1994; Sasaki et al., 2000] модулирует активность митоКАТФ. 5-ГД полностью блокирует этот кардиопротекторный эффект, подтверждая участие митоКАТФ в защите миокарда от ишемических повреждений. Следует отметить, что сами по себе глибенкламид и 5-ГД не оказывают никакого влияния [Gross and Fryer, 1999]. Неспособность специфического ингибитора цитоКАТФ, HMR-1883, ингибировать анти-ишемический эффект активаторов митоКАТФ [Fryer et al., 2000], также говорит в пользу участия в защитном механизме МХ канала. Следует отметить, что неселективные активаторы КАТФ каналов, такие как пинацидил, оказывая кардиопротекторное действие, вызывали побочный проаритмический эффект и приводили к существенному понижению давления у подопытных животных [Grover, 1997]. Подобные нежелательные последствия способны свести на нет кардиопротекторное действие данных препаратов. Использование селективных активаторов МХ или цитоплазматического КАТФ каналов позволило бы избежать таких побочных явлений. Можно ожидать, что селективные активаторы цитоКАТФ, такие как HMR-1883, оказывая антиаритмическое действие, не будут негативно влиять на ишемическую адаптацию, вызванную активацией митоКАТФ. Так, показано, что HMR-1883 является потенциальным антифибрилляторным агентом при острой ишемии на модели собаки [Billman et al., 1998]. И наоборот, сайт-специфичные активаторы митоКАТФ не будут оказывать проаритмического действия [Gross and Fryer, 1999]. 2.2.2 Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ Несмотря на то, что роль митоКАТФ в адаптации организма к гипоксии доказана, остается неясным, каким образом открытие канала приводит к защите сердца от ишемических повреждений. Недавно были предложены три различных механизма защиты сердца при гипоксии, которые могут проявляться при открытии митоКАТФ: 1) механизм, при котором усиление входа К+ в МХ ведет к набуханию матрикса, что предупреждает распад АТФ; 2) механизм, основанный на снижении концентрации Са2+ в МХ, что уменьшает вероятность открытия митохондриальной поры и предупреждает гибель кардиомиоцитов; 3) механизм, связанный с изменением уровня активных форм кислорода (АФК), повышенная концентрация которых, с одной стороны, активирует ряд ферментов, а с другой, необратимо повреждает мышечную ткань при гипоксии. Известно, что при набухании МХ усиливается окисление жирных кислот и увеличивается продукция АТФ [Grover, 2000; Halestrap, 1989]. Хотя ранее было продемонстрировано, что набухание МХ является следствием открытия митоКАТФ, вопрос о связи этого эффекта непосредственно с сохранением целостности МХ после ишемии и реперфузии остается открытым [Lim et al., 2002]. Идея о том, что потребление К+ митохондриями и сопутствующее ему набухание этих органелл оказывает кардиопротекторный эффект, подтверждается тем, что активация других митохондриальных К+ каналов, например, Ca2+-активируемого К+ канала, также предупреждает ишемические повреждения [Xu et al., 2002]. Однако, этот эффект не блокируется ингибитором митоКАТФ, что наблюдается при «preconditioning». Следовательно, именно митоКАТФ задействован в формировании феномена «preconditioning». |
Copyright © 2012 г.
При использовании материалов - ссылка на сайт обязательна.