 |
|
МЕНЮ
|
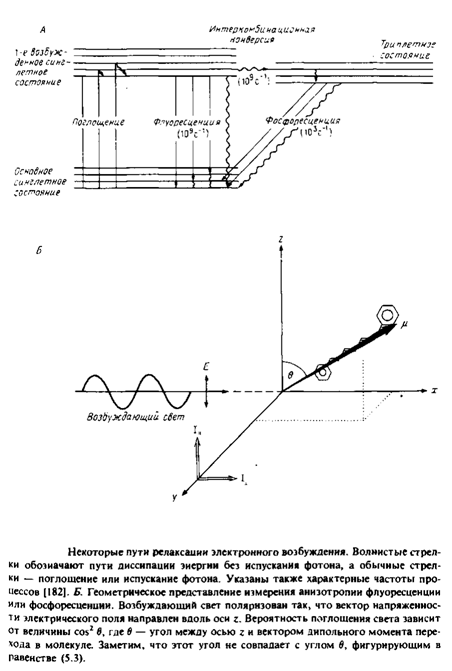
Реферат: Динамическое поведение мембранных систем и липидно-белковые взаимодействияРис. 4 иллюстрирует принципы, лежащие в основе применения флуоресценции и фосфоресценции для анализа вращения молекул. Молекула, поглотившая фотон, за время 10" 11 с переходит в возбужденное синглетное состояние Si. Термин «синглетное» указывает на то, что спин электрона при поглощении фотона не изменяется. Обычно возвращение в основное, невозбужденное состояние происходит за время - 10"8 с. Причинами такого перехода являются столкновение с соседними молекулами, диссипация энергии в виде тепла или испускание фотона. Характерное время перехода популяции молекул из синглетного возбужденного состояния в основное равно тт. Некоторые молекулы с большой вероятностью переходят из синглетного возбужденного состояния в триплетное, и этот переход сопровождается изменением спина электрона. Испускание фотона при переходе молекулы из этого состояния в основное называется фосфоресценцией. Время жизни молекулы в триплетном состоянии относительно велико, что связано с изменением спинового состояния электрона при испускании фотона. Рассмотрим принцип метода деполяризации. Свет, поляризованный вдоль оси z, будет предпочтительно возбуждать молекулы, у которых дипольный момент перехода имеет такую же ориентацию. Вероятность возбуждения пропорциональна cos2 в, где в — угол между дипольиым моментом перехода и вектором напряженности электрического поля. Такая же зависимость от cos2 в имеет место и для процесса испускания фотона, поэтому молекула, у которой дипольный момент перехода ориентирован вдоль оси z, будет испускать свет с такой же поляризацией. Испускаемый свет анализируют с помощью поляризаторов, определяя интенсивность компоненты, параллельной и перпендикулярной направлению поляризации возбуждающего света.
Анизотропия г связана с /, и IL следующим образом:
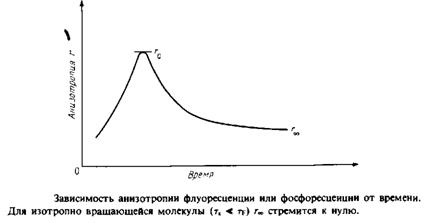
Если между моментами поглощения и испускания света молекула не вращается, то /, будет больше, чем 1±, и г будет иметь максимальное значение 0.4. Если же молекулы, находясь в возбужденном состоянии, совершают интенсивные изотропные вращательные движения, то г будет равно нулю, поскольку в среднем молекулы к моменту испускания света будут ориентированы беспорядочно. При тк = tf величина г будет зависеть от характера движения. При этом т ~ 1 и ту ' отражают скорости вращения и испускания света. Для того чтобы разграничить динамические эффекты и эффекты, связанные с ограничениями, налагаемыми на диапазон допустимых движений, определяют временную зависимость анизотропии r света, испускаемого молекулой после возбуждения. Значение г в данный момент времени отражает степень разу-порядочивания молекул относительно вектора поляризации за вре- ■ мя, прошедшее после поглощения возбуждающего импульса света. Спад r от максимального значения го задается выражением
тк — время вращательной корреляции, оно определяется скоростью затухания. Если при / оо все молекулы окажутся беспорядочно ориентированными, то л в пределе будет равно нулю. Однако если движение молекул ограничено, то их ориентация никогда не будет совершенно случайной. В такой ситуации г» не будет равно нулю, и с помощью этого параметра можно определить степень ограничения, которое налагается на подвижность зонда его окружением. Параметр упорядоченности определяется из этих величин так же, как в случае ЭПР или ЯМР:
К сожалению, такие кинетические измерения проводятся редко, большинство экспериментов выполнено в стационарном режиме при непрерывном возбуждении и испускании. Значение г, полученное таким способом, носит усредненный характер и определяется как
Рассмотрим следующие три случая. 1. Очень быстрое движение: 7"= г„, предельному значению. 2. Очень медленное движение или его отсутствие: г = го, максимальному значению. 3. Случаи, представляющие практический интерес '- 7 зависит как от скорости движений, так и от ограничений, налагаемых на них. Без соответствующей корректировки определение микровязкости по результатам измерений 7 не будет сводиться к простым динамическим измерениям. Вязкость бислоя весьма высока, поэтому время вращательной корреляции небольших флуоресцентных и ЭПР-зондов составляет 10~8—10 с. В воде, имеющей вязкость 0,01 пуаз, молекулы такого размера вращались бы по крайней мере в 100 раз быстрее. Мембранные белки имеют значительно большие размеры, чем упомянутые метки, и поэтому вращаются гораздо медленнее. Чтобы метки, «пришитые» к этим белкам, были чувствительны к их вращению, время жизни меток в возбужденном состоянии должно составлять порядка 10~3 с. 3. Вращение мембранных белков Коэффициент вращательной диффузии мембранного белка, находящегося в плоскости бислоя, D., можно найти, представив белко вую молекулу в виде цилиндра, который вращается вокрут одной оси. Пусть мембрана имеет вязкость г) и толщину h, а радиус цилиндра равен а. Тогда
Часто наряду с фу применяется время вращательной релаксации ф, = 1/. Для белковой молекулы радиусом 25 А, находящейся в мембране толщиной 40 А и вязкостью 5 пуаз, величина ф, по оценкам составляет около 35 мкс. С количественной точки зрения это уравнение, описывающее вращение белка в бислое, не вполне строго, но зависимость времени вращательной релаксации от эффективного радиуса вращающейся белковой молекулы сомнений не вызывает. Это оказалось весьма полезным для исследования процессов агрегации белков внутри мембраны. Методы, применяемые для изучения вращения белков в бислое, должны быть способны регистрировать времена вращения от 10"5 до 10 ~3 с. Обычный метод измерения деполяризации флуоресценции в этом случае непригоден, поскольку время жизни молекул в возбужденном состоянии составляет около 10"8 с, и в таком временном масштабе молекулы белков представляются неподвижными. Успешно использовались три метода. 1. К исследуемому белку присоединяют зонд, время жизни которого в возбужденном триплетном состоянии достаточно велико. Если метка жестко связана с белком, то для регистрации вращения белка можно использовать измерение анизотропии фосфоресценции. Для таких измерений оказались пригодными производные эозина, поскольку время жизни эозина в триплетном состоянии составляет примерно 2 мс. В основе этого подхода лежат те же принципы, что и в основе метода деполяризации флуоресценции, но используется другая молекулярная модель движения, вызывающего деполяризацию. Эксперимент состоит в определении характерного времени затухания анизотропии фосфоресценции. Если система гетерогенна, могут возникнуть определенные трудности, связанные с количественным анализом экспоненциальной кривой затухания анизотропии. Проблемы возникают и в тех случаях, если метка может локально вращаться на поверхности мембранного белка или если у белковой молекулы имеются гибкие сегменты. Те же метки могут использоваться и для анализа вращения белков с использованием временной зависимости дихроизма поглощения. В этом случае изменения в ориентации дипольного момента перехода регистрируются по поглощению света, поляризованного параллельно и перпендикулярно оси поляризации начального импульса, используемого для возбуждения молекул. 2. Известны случаи, когда сами молекулы белка содержат группы, переходящие при флеш-фотолизе в долгоживущее возбужденное состояние, параметры которого можно оценить с помощью дихроизма поглощения. В качестве примера можно привести родопсин и бактериородопсин, где используются возбужденное состояние связанного ретиналя и возбужденные состояния, наблюдающиеся при фотолизе комплексов цитохром—СО с использованием цитохром с-оксидазы и цитохрома Р450. Измерения можно проводить in situ или с очищенным белком, встроенным в фосфолипидные везикулы. 3. С помощью обычной ЭПР-спектроскопии не удается регистрировать вращения, характерная частота которых равна частоте вращения мембранных белков. Однако разработан специальный метод — ЭПР с переносом насыщения, диапазон чувствительности которого очень широк — от 10"7 до 10 ~J с. Этот метод применялся при изучении вращения нескольких мембранных белков с ковалентно пришитыми к ним спиновыми метками. Недостаток метода состоит в том, что в случае анизотропного молекулярного движения спектры с трудом поддаются интерпретации. 3.1 Примеры вращения белков Внутримембранные белки характеризуются широким спектром времен вращательной релаксации. На одном конце временной шкалы находится родопсин, который, по-видимому, свободно вращается в мембране наружного сегмента палочки сетчатки, а на другом — бактериородописин, который образует в пурпурной мембране упорядоченную кристаллическую решетку и неподвижен. Для нескольких очищенных белков, встроенных в фосфолипидные везикулы, зависит от концентрации. Это позволяет предположить, что при уменьшении соотношения липид/белок происходит самоагрегация этих белков. Такая картина характерна для бактериородопсина, цитохром с-оксидазы, белка полосы 3, Са2 + -АТРазы и цитохрома Р450. Возможно, именно с самоагрегацией связана гетерогенность, наблюдаемая, в частности, для белка полосы 3 в тенях эритроцитов и Са2 + -АТРазы из саркоплазматического ретикулума. Вообще говоря, мембранные белки, по-видимому, вращаются в плоскости мембраны и скорость вращения согласуется с величиной, ожидаемой исходя из простой гидродинамической модели. Дополнение. Вращение белка полосы 3 и цитохрома Р450: выявление межмолекулярных взаимодействий Из двух указанных белков — белка полосы 3 и цитохрома Р450 — наибольший интерес представляет первый, поскольку по имеющимся биохимическим данным можно предположить, что этот анионный переносчик связан со спектрино-актиновой сетью цитоскелета. Обнаружение того факта, что значительная часть белковых молекул способна к быстрому вращению, послужило основанием для создания моделей, согласно которым белок полосы 3 иммобилизован не полностью или образует со спектрином временную связь. Однако ситуация осложнялась тем, что при лизисе клеток, необходимом для получения теней эритроцитов, комплекс с цитоскелетом может разрушаться. В интактной клетке белок полосы 3 совершает только медленные вращательные движения с характерным временем 0,1 — 1 мс. Это больше согласуется с отсутствием поступательного движения белка полосы 3, хотя физическая причина такого ограничения подвижности не установлена. Цитохром Р450 акцептирует электроны от цитохром Р450-редук-тазы в электронтранспортной цепи микросом. Многих ученых интересовал вопрос: образуют ли эти молекулы долгоживущий комплекс в мембране или их взаимодействие сводится к простым столкновениям в результате диффузии? При низком соотношении липид/белок цитохром Р450 агрегирует и вращение его замедляется. Однако в присутствии стехиометрических количеств редуктазы цитохром Р450 в сходных условиях остается мобильным. Это свидетельствует о взаимодействии двух указанных белков и образовании комплекса между ними со стехиометрией 1:1, которое препятствует агрегации Р450. 4. Латеральная диффузия липидов и белков в мембранах Способность мембранных компонентов к латеральной диффузии — это одно из очевидных условий, принятых в жидкостно-мозаичной модели. Для измерения коэффициента латеральной диффузии применялось несколько методов. Рассмотрим три из них.
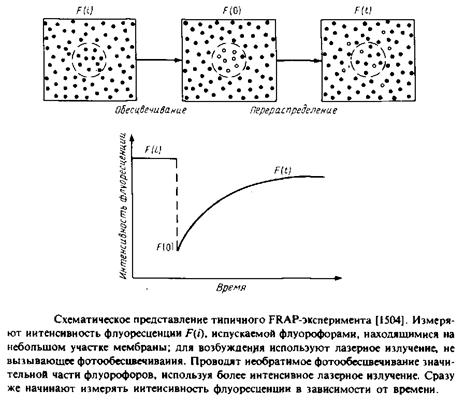
1.Осуществляют слияние клеток, несущих на своей поверхности различные маркеры, с образованием гетерокарионов. После слияния оценивают скорость перераспределения поверхностных маркеров с помощью флуоресцентной микроскопии и с использованием меченных различными флуоресцентными метками антител против специфических поверхностных антигенов. 2.Осуществляют перераспределение мембранных белков с помощью электрофореза in situ. При внесении клеток в электрическое поле белки концентрируются в одной части мембраны, и коэффициент латеральной диффузии определяют по данным о скорости их перераспределения после отключения электрического поля. За перераспределением белков можно наблюдать с помощью электронной микроскопии или флуоресцентных методов. 3.Чаще всего используется метод, основанный на восстановлении способности белков к флуоресценции после фотообесцвечивания. Суть метода представлена на рис. 5.6. С помощью мощного лазерного луча обесцвечивают пятно диаметром - 1 мкм на равномерно меченной с помощью флуоресцентной метки мембране. Далее исследуют кинетику разгорания флуоресценции этого участка и по скорости восстановления флуоресценции прямо оценивают скорость латеральной диффузии флуоресцентных зондов на обесцвеченный участок из областей мембраны, прилегающих к нему. С помощью этого метода можно исследовать амфифильные флуоресцентные зонды, меченые фосфолипиды и белки, несущие флуоресцентные метки. Структурные формулы некоторых наиболее часто используемых зондов приведены в табл. 5.1. Можно ожидать, что применение видеотехники для изучения перераспределения флуоресцентных молекул в мембране расширит рамки применения этого метода. Использование двух первых методов сопровождается значительными нарушениями клеток или мембран. Метод FRAP является более щадящим. Опасения, связанные с возможностью повреждений клеток лазерным лучом, были сняты. Метод можно также применять для исследования модельных мембранных бислоев и монослоев, а также интактных клеток или фрагментов биомембран. Фокусируя лазерный луч диаметром ~ 1 мкм в разных точках, можно наблюдать за датеральной диффузией мембранных компонентов в различных участках мембраны. Коэффициенты поступательной диффузии, которые можно определить с помощью этого метода, варьируют от Ю-7 до 10 ~12 см2/с. Коэффициент диффузии 10 "12 см2/с в рамках этого подхода соответствует отсутствию движения. Значение D = 10 "8 см2/с, характерное для липидов в биомембранах, отвечает диффузии на расстояние около 2 мкм за 1 с. 4.1 Модели, описывающие латеральную диффузию Для описания поступательной диффузии мембранных белков часто применяется гидродинамическая модель Саффмана и Дельбрюка. В этой модели рассматривается диффузия белка в тонком вязком слое; при этом считается, что растворитель является сплошной средой, т. е. молекулы растворителя малы по сравнению с диффундирующими молекулами. В рамках этой модели выполняется следующее соотношение:
где т)м и 1), — вязкости мембранной и водной фаз соответственно, а — радиус белковой молекулы, имеющей форму цилиндра, h — толщина мембраны. Из модели следует, что скорость поступательной диффузии слабо зависит от размера молекулы, что подтверждается полученными к настоящему времени экспериментальными данными.
Наконец, несколько подходов использовалось для теоретического объяснения влияния мембранных белков на латеральную диффузию белков и липидов в мембране. Дело в том, что белки оказываются на пути диффундирующих молекул и вынуждают их перемещаться только в пространстве, не занятом белками. Обычно из-за этого коэффициенты поступательной диффузии в биомембранах оказываются примерно в 10 раз меньше, чем в чисто липидных бислоях. 4.2 Примеры латеральной диффузии компонентов мембран Коэффициенты поступательной диффузии некоторых мембранных липидов и белков приведены в табл. 5.2. Коэффициенты диффузии флуоресцентных меток и аналогов липидов в фосфолипидных мультибислоях или в крупных однослойных везикулах равен 10"8 см2/с. При изучении поведения этих же зондов в разнообразных природных мембранах коэффициент диффузии обычно оказывается примерно в 10 раз меньше. Этот феномен, как правило, объясняют присутствием в биомембранах белков, препятствующих латеральной диффузии. Скорость латеральной диффузии фосфолипидов с разными полярными головками различается слабо, однако в реконструированных везикулах гангли-озиды диффундируют медленнее. При переходе мембранного бислоя в состояние геля скорость латеральной диффузии уменьшается более чем на два порядка {D < 10" 10 см2/с). В жидком бислое модельных мембран и в биомембранах липиды обычно диффундируют свободно, хотя имеются редкие исключения. Совсем иначе обстоит дело с латеральной диффузией мембранных белков. Исследование поведения нескольких белков в реконструированных везикулах, пока не позволяющее сделать окончательные выводы, тем не менее указывает на то, что коэффициенты поступательной диффузии белков разных размеров очень близки и что белки диффундируют лишь немного медленее, чем липиды. Исследованные белки сильно различались по числу трансмембранных сегментов и по степени ассоциированности, и тем не менее диффундировали с одинаковой скоростью. Модель Саффмана—Дельбрюка предсказывает очень слабую зависимость скорости диффузии от размеров молекул. В реконструированных везикулах с высоким содержанием белка могут происходить агрегация последнего и латеральное разделение фаз, приводящие к иммобилизации части белка. В биомембранах коэффициенты латеральной диффузии белков обычно в 100—1000 раз меньше, чем в модельных системах с низкой концентрацией белка. Например, родопсин, по-видимому, свободно диффундирует в биомембране— D = 4-10"9 см2/с. Поскольку мембраны, содержащие родопсин, характеризуются высоким отношением белок/липид, ясно, что обычно наблюдаемая медленная диффузия других мембранных белков не может быть обусловлена исключительно присутствием белков в бислое. Есть и еще один интересный факт: восстановление флуоресценции в экспериментах FRAP происходит не полностью и нередко составляет менее 75%. Это означает, что часть изучаемой популяции белков ^ неподвижна. Вопрос о том, что ограничивает латеральную диффузию мембранных белков, имеет ключевое значение. Маловероятно, что это обусловлено агрегацией мембранных белков, поскольку для объяснения большинства данных потребовалось бы предположить, что образуются слишком крупные агрегаты. Одно из возможных объяснений состоит в том, что мембранные белки,малоподвижны, поскольку связаны с внеклеточным матриксом или с ци-тоскелетом. Обнаружилось, что, оказывая воздействие на внеклеточный матрикс, можно влиять на латеральную диффузию мембранных белков, однако имеются и другие данные, согласно которым эта связь не столь уж важна. |
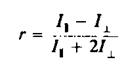
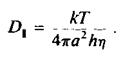
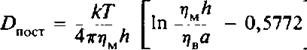

Copyright © 2012 г.
При использовании материалов - ссылка на сайт обязательна.